Acrocanthosaurus
.jpg)
Acrocanthosaurus (meaning “high-spined lizard”) is a genus of theropod dinosaur that existed in what is now North America during the Aptian and early Albian stages of the Early Cretaceous. Like most dinosaur genera, Acrocanthosaurus contains only a single species, A. atokensis. Its fossil remains are found mainly in the U.S. states of Oklahoma, Texas, and Wyoming, although teeth attributed to Acrocanthosaurus have been found as far east as Maryland.
Acrocanthosaurus was a bipedal predator. As the name suggests, it is best known for the high neural spines on many of its vertebrae, which most likely supported a ridge of muscle over the animal’s neck, back and hips. Acrocanthosaurus was one of the largest theropods, reaching 11.5 m (38 ft) in length, and weighing up to 6.2 tonnes (6.8 short tons). Large theropod footprints discovered in Texas may have been made by Acrocanthosaurus, although there is no direct association with skeletal remains.
Recent discoveries have elucidated many details of its anatomy, allowing for specialized studies focusing on its brain structure and forelimb function. Acrocanthosaurus was the largest theropod in its ecosystem and likely an apex predator which preyed on sauropods, ornithopods, and ankylosaurs.

Acrocanthosaurus was big for its time, a family trait that is shared with some other carcharodontosaurids such as Carcharodontosaurus and Giganotosaurus. At eleven and a half meters, Acrocanthosaurus would have been the largest predator of its time and locale. Its diet as a result probably consisted of hadrosaurs and smaller sauropods, dinosaurs that were large enough to provide sufficient sustenance, while being too slow to escape. Study of the area that the main Acrocanthosaurus remains come from suggest that it was probably the apex predator of its location, with most other predators such as Deinonychus being much smaller.
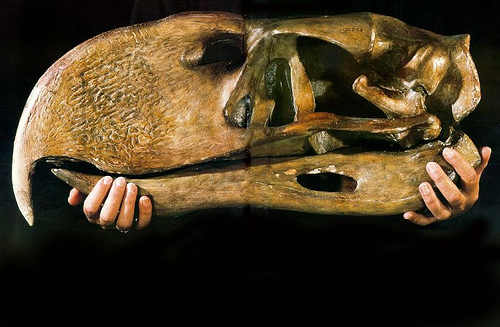
The skull of Acrocanthosaurus featured large fenestra, a necessary adaptation to reduce the weight of its huge skull that could approach up to a one hundred and thirty centimeters long. The teeth of Acrocanthosaurus were curved and serrated like other members of the carcharodontosaurid group. The maxilla and premaxilla contained a total of around thirty-eight teeth. The teeth in the lower jaw are generally smaller than those above and can approach up to thirty in number. Another carcharodontosaurid trait is the bony brow ridge above the eye, formed by the lacrimal and postorbital bones coming together.

Computer reconstruction of the inner ear has shown that the ‘resting position’ of the head of Acrocanthosaurus was twenty-five degrees below zero horizontal. This may give the impression that Acrocanthosaurus usually walked around looking slightly towards the ground.
Reconstruction of an Acrocanthosaurus forelimb suggests that there would have been large amounts of cartilage between the bones. This comes from the fact that bones themselves do not make perfect joints and would need the extra cartilage in order to articulate properly. The arms of Acrocanthosaurus did not have a huge range of motions. The arm could not fully extend and could only manage limited flexing. The humerus could retract back quite away, as if Acrocanthosaurus was pulling something towards its chest. As is commonly seen in larger theropods, the fore arm could not twist like a human arm can. When at rest the arms would have faced medially inwards, like when you clap your hands together. Acrocanthosaurus had three digits on the end of its arms with the first and second claws probably being permanently flexed. The third and smallest claw may have been able to retract as well.
Altogether, Acrocanthosaurus may have grabbed large prey such as sauropods like Paluxysaurus with its jaws and then latched onto it with its claws. The neck vertebrae also interlocked together for greater rigidity which means that Acrocanthosauruscould hold onto large prey with its jaws without sustaining injury to the neck. Lighter prey such as ornithopod dinosaurs like Tenontosaurus may have been pulled towards Acrocanthosaurus while it continued to work with its jaws, whereas it would probably have to pull itself onto heavier prey. Alternatively it may have held its prey with its jaws while repeatedly slashing at it with its claws, the more the prey struggled, the worse its wounds became.

One usual feature of Acrocanthosaurus is the large neural spines on the vertebrae of the neck, back, hips and upper tail. It is not thought that it had a sail on its back like Spinosaurus (which was unrelated, and had much larger neural spines), but rather it is thought the spines had attachments for powerful muscles, similar to those found in modern bison. Its not entirely clear what the purpose of these spines and muscles were – possibilities include fat storage, communication, or temperature regulation.
Acrocanthosaurus was a carnivore, but is not believed to have been a fast runner. Its forelimbs and shoulders are also unusual, and seem to have been very strong, had lots of cartilage, but been quite stiff with very limited movement. It is thought that the forelimbs hung down and inwards, and would not have been used for seizing prey. Acrocanthosaurus may instead have seized prey with its jaws, and used its forelimbs to prevent the prey escaping. It is also possible that Acrocanthosaurus may have held the prey in its jaws, and used the claws in its forelimbs to tear gashes into the prey.