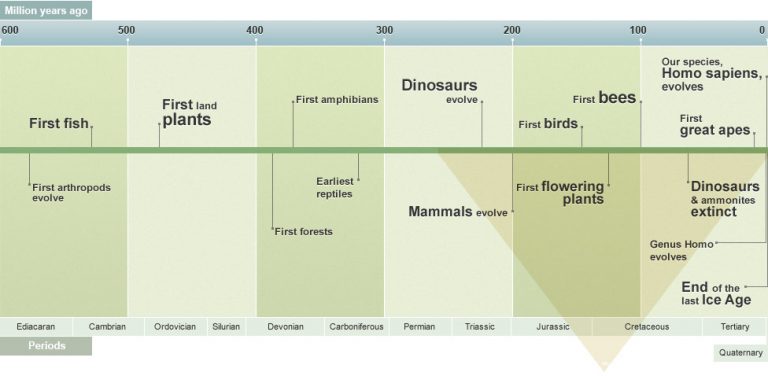
The Cretaceous is a geologic period and system that spans 79 million years from the end of the Jurassic Period 145 million years ago (Mya) to the beginning of the Paleogene Period 66 Mya. It is the last period of the Mesozoic Era. The Cretaceous Period is usually abbreviated K, for its German translation Kreide (chalk).
The Cretaceous was a period with a relatively warm climate, resulting in high eustatic sea levels that created numerous shallow inland seas. These oceans and seas were populated with now-extinct marine reptiles, ammonites and rudists, while dinosaurs continued to dominate on land. During this time, new groups of mammals and birds, as well as flowering plants, appeared. The Cretaceous ended with a large mass extinction, the Cretaceous–Paleogene extinction event, in which many groups, including non-avian dinosaurs, pterosaurs and large marine reptiles died out. The end of the Cretaceous is defined by the abrupt Cretaceous–Paleogene boundary (K–Pg boundary), a geologic signature associated with the mass extinction which lies between the Mesozoic and Cenozoic eras.
Research history
The Cretaceous as a separate period was first defined by Belgian geologist Jean d’Omalius d’Halloy in 1822, using strata in the Paris Basin and named for the extensive beds of chalk (calcium carbonate deposited by the shells of marine invertebrates, principally coccoliths), found in the upper Cretaceous of Western Europe. The name Cretaceous was derived from Latin creta, meaning chalk.
Stratigraphic subdivisions
The Cretaceous is divided into Early and Late Cretaceous epochs, or Lower and Upper Cretaceous series. In older literature the Cretaceous is sometimes divided into three series: Neocomian (lower/early), Gallic (middle) and Senonian (upper/late). A subdivision in eleven stages, all originating from European stratigraphy, is now used worldwide. In many parts of the world, alternative local subdivisions are still in use.
As with other older geologic periods, the rock beds of the Cretaceous are well identified but the exact age of the system’s base is uncertain by a few million years. No great extinction or burst of diversity separates the Cretaceous from the Jurassic. However, the top of the system is sharply defined, being placed at an iridium-rich layer found worldwide that is believed to be associated with the Chicxulub impact crater, with its boundaries circumscribing parts of the Yucatán Peninsula and into the Gulf of Mexico. This layer has been dated at 66.043 Ma.
A 140 Ma age for the Jurassic-Cretaceous boundary instead of the usually accepted 145 Ma was proposed in 2014 based on a stratigraphic study of Vaca Muerta Formation in Neuquén Basin, Argentina. Víctor Ramos, one of the authors of the study proposing the 140 Ma boundary age sees the study as a “first step” toward formally changing the age in the International Union of Geological Sciences.
From youngest to oldest, the subdivisions of the Cretaceous period are:
Late Cretaceous
Maastrichtian – (66-72.1 MYA)
Campanian – (72.1-83.6 MYA)
Santonian – (83.6-86.3 MYA)
Coniacian – (86.3-89.8 MYA)
Turonian – (89.8-93.9 MYA)
Cenomanian – (93.9-100.5 MYA)
Early Cretaceous
Albian – (100.5-113.0 MYA)
Aptian – (113.0-125.0 MYA)
Barremian – (125.0-129.4 MYA)
Hauterivian – (129.4-132.9 MYA)
Valanginian – (132.9-139.8 MYA)
Berriasian – (139.8-145.0 MYA)
Rock formations
The high sea level and warm climate of the Cretaceous meant large areas of the continents were covered by warm, shallow seas, providing habitat for many marine organisms. The Cretaceous was named for the extensive chalk deposits of this age in Europe, but in many parts of the world, the deposits from the Cretaceous are of marine limestone, a rock type that is formed under warm, shallow marine circumstances. Due to the high sea level there was extensive space for such sedimentation. Because of the relatively young age and great thickness of the system, Cretaceous rocks are evident in many areas worldwide.
Chalk is a rock type characteristic for (but not restricted to) the Cretaceous. It consists of coccoliths, microscopically small calcite skeletons of coccolithophores, a type of algae that prospered in the Cretaceous seas.
In northwestern Europe, chalk deposits from the Upper Cretaceous are characteristic for the Chalk Group, which forms the white cliffs of Dover on the south coast of England and similar cliffs on the French Normandian coast. The group is found in England, northern France, the low countries, northern Germany, Denmark and in the subsurface of the southern part of the North Sea. Chalk is not easily consolidated and the Chalk Group still consists of loose sediments in many places. The group also has other limestones and arenites. Among the fossils it contains are sea urchins, belemnites, ammonites and sea reptiles such as Mosasaurus.
In southern Europe, the Cretaceous is usually a marine system consisting of competent limestone beds or incompetent marls. Because the Alpine mountain chains did not yet exist in the Cretaceous, these deposits formed on the southern edge of the European continental shelf, at the margin of the Tethys Ocean.
Stagnation of deep sea currents in middle Cretaceous times caused anoxic conditions in the sea water leaving the deposited organic matter undecomposed. Half the worlds petroleum reserves were laid down at this time in the anoxic conditions of what would become the Persian Gulf and Gulf of Mexico. In many places around the world, dark anoxic shales were formed during this interval. These shales are an important source rock for oil and gas, for example in the subsurface of the North Sea.
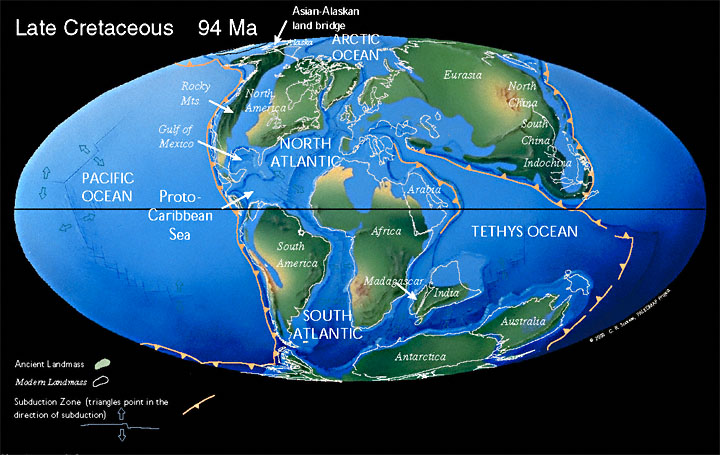
During the Cretaceous, the late-Paleozoic-to-early-Mesozoic supercontinent of Pangaea completed its tectonic breakup into the present-day continents, although their positions were substantially different at the time. As the Atlantic Ocean widened, the convergent-margin mountain building (orogenies) that had begun during the Jurassic continued in the North American Cordillera, as the Nevadan orogeny was followed by the Sevier and Laramide orogenies.
Though Gondwana was still intact in the beginning of the Cretaceous, it broke up as South America, Antarctica and Australia rifted away from Africa (though India and Madagascar remained attached to each other); thus, the South Atlantic and Indian Oceans were newly formed. Such active rifting lifted great undersea mountain chains along the welts, raising eustatic sea levels worldwide. To the north of Africa the Tethys Sea continued to narrow. Broad shallow seas advanced across central North America (the Western Interior Seaway) and Europe, then receded late in the period, leaving thick marine deposits sandwiched between coal beds. At the peak of the Cretaceous transgression, one-third of Earth’s present land area was submerged.
Climate
The cooling trend of last epoch of the Jurassic continued into the first age of the Cretaceous. There is evidence that snowfalls were common in the higher latitudes and the tropics became wetter than during the Triassic and Jurassic. Glaciation was however restricted to high-latitude mountains, though seasonal snow may have existed farther from the poles. Rafting by ice of stones into marine environments occurred during much of the Cretaceous but evidence of deposition directly from glaciers is limited to the Early Cretaceous of the Eromanga Basin in southern Australia.
A very gentle temperature gradient from the equator to the poles meant weaker global winds, which drive the ocean currents, resulted in less upwelling and more stagnant oceans than today. This is evidenced by widespread black shale deposition and frequent anoxic events. Sediment cores show that tropical sea surface temperatures may have briefly been as warm as 42 °C (108 °F), 17 °C (31 °F) warmer than at present, and that they averaged around 37 °C (99 °F). Meanwhile, deep ocean temperatures were as much as 15 to 20 °C (27 to 36 °F) warmer than today’s.
Flora
Flowering plants (angiosperms) spread during this period, although they did not become predominant until the Campanian Age near the end of the period. Their evolution was aided by the appearance of bees; in fact angiosperms and insects are a good example of coevolution. The first representatives of many leafy trees, including figs, planes and magnolias, appeared in the Cretaceous. At the same time, some earlier Mesozoic gymnosperms continued to thrive; pehuéns (monkey puzzle trees, Araucaria) and other conifers being notably plentiful and widespread. Some fern orders such as Gleicheniales appeared as early in the fossil record as the Cretaceous, and achieved an early broad distribution. Gymnosperm taxa like Bennettitales and hirmerellan conifers died out before the end of the period.
Terrestrial fauna
On land, mammals were generally small sized, but a very relevant component of the fauna, with cimolodont multituberculates outnumbering dinosaurs in some sites. Neither true marsupials nor placentals existed until the very end, but a variety of non-marsupial metatherians and non-placental eutherians had already began to diversify greatly, ranging as carnivores (Deltatheroida), aquatic foragers (Stagodontidae) and herbivores (Schowalteria, Zhelestidae). Various “archaic” groups like eutriconodonts were common in the Early Cretaceous, but by the Late Cretaceous northern mammalian faunas were dominated by multituberculates and therians, with dryolestoids dominating South America.
The apex predators were archosaurian reptiles, especially dinosaurs, which were at their most diverse stage. Pterosaurs were common in the early and middle Cretaceous, but as the Cretaceous proceeded they declined for poorly understood reasons (once thought to be due to competition with early birds, but now it is understood avian adaptive radiation is not consistent with pterosaur decline), and by the end of the period only two highly specialized families remained.
Insects diversified during the Cretaceous, and the oldest known ants, termites and some lepidopterans, akin to butterflies and moths, appeared. Aphids, grasshoppers and gall wasps appeared.
Marine fauna
In the seas, rays, modern sharks and teleosts became common. Marine reptiles included ichthyosaurs in the early and mid-Cretaceous (becoming extinct during the late Cretaceous Cenomanian-Turonian anoxic event), plesiosaurs throughout the entire period, and mosasaurs appearing in the Late Cretaceous.
Baculites, an ammonite genus with a straight shell, flourished in the seas along with reef-building rudist clams. The Hesperornithiformes were flightless, marine diving birds that swam like grebes. Globotruncanid Foraminifera and echinoderms such as sea urchins and starfish (sea stars) thrived. The first radiation of the diatoms (generally siliceous shelled, rather than calcareous) in the oceans occurred during the Cretaceous; freshwater diatoms did not appear until the Miocene. The Cretaceous was also an important interval in the evolution of bioerosion, the production of borings and scrapings in rocks, hardgrounds and shells.
End-Cretaceous extinction event
The impact of a large body with the Earth may have been the punctuation mark at the end of a progressive decline in biodiversity during the Maastrichtian Age of the Cretaceous Period. The result was the extinction of three-quarters of Earth’s plant and animal species. The impact created the sharp break known as K–Pg boundary (formerly known as the K–T boundary). Earth’s biodiversity required substantial time to recover from this event, despite the probable existence of an abundance of vacant ecological niches.
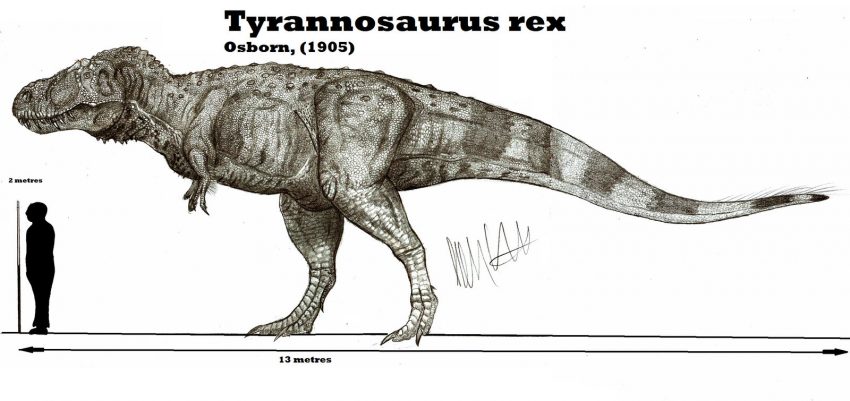
Despite the severity of K-Pg extinction event, there was significant variability in the rate of extinction between and within different clades. Species which depended on photosynthesis declined or became extinct as atmospheric particles blocked solar energy. As is the case today, photosynthesizing organisms, such as phytoplankton and land plants, formed the primary part of the food chain in the late Cretaceous, and all else that depended on them suffered as well. Herbivorous animals, which depended on plants and plankton as their food, died out as their food sources became scarce; consequently, the top predators such as Tyrannosaurus rex also perished. Yet only three major groups of tetrapods disappeared completely; the non-avian dinosaurs, the plesiosaurs and the pterosaurs. The other Cretaceous groups that did not survive into the Cenozoic era, the ichthyosaurs and last remaining temnospondyls and non-mammalian cynodonts were already extinct millions of years before the event occurred.
Coccolithophorids and molluscs, including ammonites, rudists, freshwater snails and mussels, as well as organisms whose food chain included these shell builders, became extinct or suffered heavy losses. For example, it is thought that ammonites were the principal food of mosasaurs, a group of giant marine reptiles that became extinct at the boundary.
Omnivores, insectivores and carrion-eaters survived the extinction event, perhaps because of the increased availability of their food sources. At the end of the Cretaceous there seem to have been no purely herbivorous or carnivorous mammals. Mammals and birds which survived the extinction fed on insects, larvae, worms and snails, which in turn fed on dead plant and animal matter. Scientists theorise that these organisms survived the collapse of plant-based food chains because they fed on detritus.
In stream communities, few groups of animals became extinct. Stream communities rely less on food from living plants and more on detritus that washes in from land. This particular ecological niche buffered them from extinction. Similar, but more complex patterns have been found in the oceans. Extinction was more severe among animals living in the water column, than among animals living on or in the sea floor. Animals in the water column are almost entirely dependent on primary production from living phytoplankton, while animals living on or in the ocean floor feed on detritus or can switch to detritus feeding.
The largest air-breathing survivors of the event, crocodilians and champsosaurs, were semi-aquatic and had access to detritus. Modern crocodilians can live as scavengers and can survive for months without food and go into hibernation when conditions are unfavourable, and their young are small, grow slowly, and feed largely on invertebrates and dead organisms or fragments of organisms for their first few years. These characteristics have been linked to crocodilian survival at the end of the Cretaceous.
Source: www.Wikipedia.org, www.NatGeo.com
_____________________________________